Janus Kinase e outras vias de sinalização

JAK 2
Janus Kinase, abreviadamente chamada de JAK, é uma família de tirosina quinases não receptores, intracelulares que fazem a transdução dos sinais mediados por citoquinas pela via sinalizadora JAK-STAT.
Transdução de sinal refere-se a qualquer processo através do qual uma célula converte um tipo de sinal ou estímulo em outro
As JAKs possuem 2 domínios de transferência de fosfatos praticamente iguais. Um destes domínios ( JH1 ) exibe a actividade da quinase, enquanto o outro domínio ( JH2 ) regula negativamente a actividade da quinase do JH1.
.jpg)
Esquema da composição de JAK 2
A família Janus kinase é composta por 4 membros, nomeadamente JAK1, JAK 2, JAK 3 e TYK2.
.jpg)
Membros da família das Janus Kinases
A falta de expressão de JAK1 leva a uma deficiente, ou nula, resposta a algumas citoquinas como o γ-interferão. JAK1 e JAK2 estão envolvidas na sinalização do interferão tipo II ( γ-interferão ) enquanto que JAK1 e TYK2 estão envolvidas na sinalização do interferão tipo I. A falta de expressão de TYK2 leva a uma função defeituosa das células NK.
Uma vez que os membros das familias dos receptores das citoquinas tipo I e tipo II não possuem actividade catalítica, dependem das tirosina quinases da família JAK para fosforilar e activar as proteínas envolvidas nas vias de transdução do sinal. Os receptores são formados por um par de polipeptídeos, exibindo assim 2 domínios intracelulares de transdução do sinal
.jpg)
Estrutura de JAK e receptores tipo I e tipo II

JAK associa-se, em cada domínio intracelular, a uma região rica em prolina, adjacente à membrana celular e denominada box1/box2

Box 1
Depois do receptor se associar com a sua respectiva citoquina/ligando sofre uma transformação conformacional, de tal modo que os 2 JAKs ficam tão próximos que se fosforilam mutuamente. A autofosforilação do JAK induz uma alteração conformacional dentro do próprio JAK, possibilitando o JAK de fazer a transdução do sinal intracelular através de factores de fosforilação e activação de transcripção, os chamados factores STAT ( signal transducer and activactor of transcription ). Os STAT activados dissociam-se do receptor e formam dímeros antes de se translocarem para o núcleo celular onde vão regular a transcripção dos genes seleccionados.
Utilizam a via de sinalização do JAK/STAT várias moléculas como, por exemplo, factores colónia estimulantes ( CSF ), prolactina, hormona de crescimento e muitas citoquinas.
JAK é uma molécula de 120-140 KDa com 7 diferentes regiões denominadas homologias ( JH1 a JH7 ). Como já referimos atrás, JH1 é o domínio importante para a actividade enzimática da JAK, com propriedades típicas de tirosina quinase tais como conservar as tirosinas necessárias para a activação do JAK. A fosforilação destas duplas de tirosina leva às alterações conformacionais necessárias, na proteína JAK, para facilitar a ligação do substracto. A homologia JH2 é um domínio pseudo-quinase, estruturalmente idêntico a JH1, e essencial para a normal actividade da quinase, embora desprovida de actividade enzimática, estando envolvida na regulação da homologia JH1. As homologias JH3 e JH4 compartilham homologia com SH2 ( domínios Src-homologia-2 ). O terminal amino ( JH 5 a JH7 ) do JAK é chamado de domínio FERM, também encontrado na família FAK, e é envolvido na associação de JAK com os receptores da citoquina e/ou outras quinases

Janus Kinase ( JAK )
As janus quinases ( JAK ), proteínas de cerca de 1150 amino-ácidos com cerca de 120-130 KDa, são componentes cruciais de múltiplas vias de sinalização, que governam diversos aspectos da vida celular, nomeadamente a sobrevivência celular, sua proliferação, diferenciação e apoptose. A activação das proteínas STAT é uma importante acção das JAK quinases. Fosforilação dos receptores das citoquinas e dos factores de crescimento pode ser o principal atributo funcional das JAK quinases. A fosforilação dos receptores activados pelas JAKs pode ser, potencialmente, o factor limitante da velocidade para o culminar das cascatas de sinalização. Neste sentido verifica-se a necessária função da JAK quinase na óptima activação das cascatas Src-quinase, via de sinalização Ras-MAPK, via de sinalização PI3K-akt e sinalização STAT seguinte à interacção dos receptores citoquina/interferão com os seus ligandos. Condições patológicas foram associadas a hipoactividade das JAK quinases, como por exemplo imunodeficiência mista severa; pelo contrário, hiperactividade das JAK quinases foram associadas a tumorigénese.

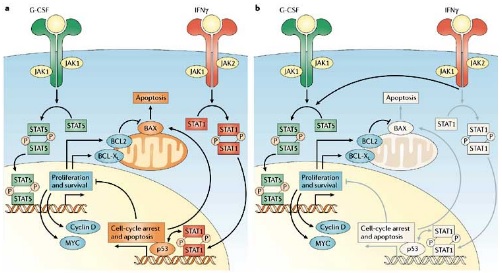
Vias de activação STAT
As JAKs controlam a sinalização promovida pela IL-2, um factor decisivo para o aumento dos linfócitos T. O bloqueio de uma JAK pode regular a resposta imune exacerbada.
A maioria dos processos de transdução de sinal, nos mamíferos, são iniciados como resultado de interacção dos receptores dos ligandos. Estas interacções resultam em alterações bioquímicas, que são processadas e enviadas para o núcleo, levando à alteração da expressão dos genes.
Contrariamente ao que apresentam os receptores dos factores de crescimento, receptores das citoquinas não possuem um domínio quinase citoplasmático.

Receptores de factor de crescimento, sinalização e transdução do sinal

Receptores de citoquina, sinalização e transdução do sinal
No entanto, foi verificado que as interacções das citoquinas com os seus receptores rapidamente induziram a fosforilação tirosina dos receptores, e de diversas outras proteínas celulares, o que sugere que estes receptores transmitem o seu sinal através das tirosina quinases celulares. A generalidade das citoquinas transmite o seu sinal por meio das JAK quinases.

Actividade tirosina quinase e acção de STAT
As PTK ( proteinas tirosina quinases convencionais ) possuem domínios catalíticos de 250-350 amino-ácidos. A localização dos domínios catalíticos quinase, na maioria das enzimas, encontra-se na terminação carboxílica da molécula, enquanto que a terminação amino apresenta um papel regulador. As PTK podem fazer parte das subfamílias Src, Tec ou de cada uma das subfamílias dos diferentes receptores de factores de crescimento, nomeadamente receptores EGF, receptores da insulina ou receptor PDGF.


Exemplo de proteína tirosina quinase : HER2
Para além dos domínios catalíticos quinases, as PTK apresentam certas outras propriedades que lhes permitem interactuar com diversos intermediários da sinalização. Estes domínios são representados por SH2 ( homologia 2 Src ), domínio SH3, PH ( domínio homologia pleckstrin ), uma tirosina com regulação negativa presente no terminal carboxílico e locais de modificação lipídica por palmitoilação ou miristilação, no terminal amino da molécula. Estes 2 últimos ( locais de modificação lipídica ) consideram-se ser importantes para a ligação das quinases à membrana.
As tirosina quinases constituintes da família JAK diferem das outras PTK pela presença de um domínio quinase adicional.
JAK 1, JAK 2 e TYK 2 estão expressas ubiquitivamente, e são codificadas por transcriptos de 5.4 Kb ( JAK 1 ), dois transcriptos de 5.3 e 5.0 Kb ( JAK 2 ) e um transcripto de 4.4 Kb ( TYK 2 ). JAK 3 é codificado por um transcripto de 4.2 Kb, que codifica uma proteína de 120 KDa, expressada predominantemente em células de origem hematopoiética. Os membros da família JAK são codificados nos cromossomas 1 ( JAK 1 ), cromossoma 9 ( JAK 2 ) e cromossoma 19 ( JAK 3 e TYK 2 )..
A estrutura única das JAK quinases distingue-as de outros membros da família PTK. O facto mais característico destas proteínas, JAK quinases, é a presença de 2 domínios ( JH 1 e JH 2 ) com extensa homologia com os domínios da tirosina quinase. Um segundo dado interessante, é a ausência de domínio SH 2 ou SH 3. Em vez disto, as JAK quinases apresentam 7 domínios denominados JAK homologias ( JH 1 a JH 7 ) que seguem a terminal amino não conservado de 30-50 amino-ácidos. Dos 2 domínios quinase identificados, apenas o JH 1 parece ser funcional, ao contrário do que parece acontecer com JH 2 que não tem certos amino-ácidos críticos, requeridos para a função quinase. As homologias JH 1 e JH 2 estão no terminal carboxílico da proteína JAK.
O domínio N-terminal das JAK, intermedeia a sua interacção com os receptores da citoquina, interferão e hormona de crescimento. A região amino terminal do JAK 1 e JAK 2 é requerida para a associação com o receptor do γ-interferão e cascata de sinalização
Domínios JH 1 e JH 2
O domínio JH 2, apesar de apresentar a maioria dos amino-ácidos característicos do domínio quinase, não tem actividade tirosina quinase, jogando, em vez disso, um papel regulador da modulação da actividade quinase de JAK 2. A deleção do domínio pseudo-quinase do JAK 2 ( JH 2 ), mas não dos domínios JH 3 a JH 7, regula negativamente a actividade catalítica JAK 2, bem como a activação STAT 5 pelo JAK 2. Verifica-se, assim, que a inibição da JAK 2 quinase é mediada por uma interacção de JH 1 e JH 2.
A autofosforilação e transfosforilação dos resíduos tirosina conservados na via de activação da JAK quinase determina os níveis da activação catalítica.
Activação da JAK quinase pelas citoquinas, interferão e hormona de crescimento
As Janus Kinase estão na sua forma inactiva nas células não estimuladas. Oligomerização dos receptores induzida por ligandos, tal como a interacção das citoquinas com o seu receptor específico, funciona como factor desencadeante do sinal de recrutamento das JAKs à proximidade dos receptores.
JAK quinases, seja como monómeros, homodímeros ou heterodímeros, têm sido envolvidas no processo de transdução do sinal iniciado por uma variedade de factores de crescimento e citoquinas.
JAK quinases estão envolvidas na sinalização por interleucinas, tais como IL-2, IL-4, IL-7, IL-9 e IL-15.
JAK 2 é activada após a estimulação pela eritropoietina do seu receptor, enquanto que JAK 1 e JAK 2 foram implicadas na sinalização por IL-3, GM-CSF e IL-5.
JAK 2 e TYK 2 são activadas por estimulação da IL-12, e considera-se jogarem, aquelas quinases, um papel crítico na mediação pelo IL-12 da diferenciação das células T.
JAK quinase joga um papel chave na via da transdução do sinal iniciada em resposta à hormona do crescimento, prolactina e G-CSF.
Também os interferãos fazem a transdução do seu sinal pelas JAK quinases, nomeadamente JAK 1, JAK 2 e TYK 2.
JAK 3 está implicada na sinalização emanada da IL-2, IL-4, IL-7, IL-15 e IL-9.

Em resumo, pode dizer-se que JAK quinases específicas, seja sózinhas ou em combinação com outras JAK quinases, podem ser preferencialmente activadas, dependendo do tipo de receptor que é activado.
Os receptores são, primariamente, substractos da activação do JAK.
Dimerização ou oligomerização dos receptores, devida à ligação aos ligandos, resulta na justaposição das JAKs que estão na vizinhança, tanto por interacções homodiméricas como heterodiméricas. Este recrutamento das JAK quinases resulta na sua fosforilação por via autofosforilação e/ou fosforilação cruzada por outras JAK quinases ou outros membros da família das tirosina quinases. Esta activação presume-se resultar num aumento da actividade da JAK quinase. As JAKs activadas fosforilarão os receptores nos locais tirosina alvo. Os locais fosfotirosina, nos receptores, podem servir de locais âncora que permitem a ligação de outros domínios SH 2, contendo moléculas sinalizadoras, tais como STAT, Src-quinases, fosfatases proteicas e outros adaptadores de proteinas sinalizadoras, como Shc, Grb 2 e Cbl.
As citoquinas, bem como a eritropoietina ou outras, têm capacidade para rapidamente induzirem a oligomerização dos receptores, levando à activação de JAK 2, com uma das subunidades dos receptores, sejam estes receptores de cadeia única ( como é o caso da eritropoietina, prolactina, hormona de crescimento e G-CSF ) ou de múltiplas cadeias ( como os receptores da IL-3, IL-5 e GM-CSF ).

O receptor associado a JAK quinase pode processar o sinal ou, subsequentemente à oligomerização do receptor, pode recrutar outras JAKs na vizinhança. Homo e heterodimerização das JAKs, seguidas pela sua activação por meios da fosforilação, resulta na propagação do sinal inicial.
O Box 1 compreende 8 resíduos de prolina, que são requeridos para a interacção de vários receptores de citoquinas ou de factores de crescimento com JAK 2. Assim a Box 1 dos receptores da prolactina, hormona de crescimento, eritropoietina, IL-6 e citoquinas associadas, que utilizam a subunidade do receptor de transdução do sinal gp130, é requerida ( a Box 1 ) para associação com JAK 2.
A estimulação de IL-2 leva à activação de JAK 3 mas não de JAK 2, enquanto que a eritropoietina transmite seu sinal por recrutar e activar a JAK 2.
As regiões Box 1 e Box 2 são regiões de gp130, e foram demonstradas serem requeridas para associação e activação de JAK 1 e JAK 2.
JAK 1 e JAK 3 constituem componentes integrais do aparelho de transdução do sinal IL-2R.
A região Box 1 tem um papel na determinação da especificidade da activação da JAK quinase.
Embora a activação de STAT seja um passo importante, concretamente para a activação da JAK quinase, vários eventos prévios à activação do STAT parecem ser únicos para as JAK quinases. Um exemplo desta interdependência, na sinalização JAK quinase, é exemplificada pelo papel das JAK quinases na sinalização dos interferãos α/β e γ. Consistente com a associação mediada pelas subunidades de múltiplos membros da família JAK, interferão tipo I ( α-interferão e β-interferão ) e tipo II ( γ-interferão ) ligam-se a 2 complexos receptores não relacionados, cada um dos quais usa JAK quinases, comuns a ambos e distintos entre si, para transmitir os seus sinais.
JAK 1, foi demonstrado ser obrigatório para iniciar a fosforilação e activação de TYK 2 num complexo receptor do interferão α/β. JAK 2 serve como quinase iniciadora que fosforila JAK 1 como parte da cascata de sinalização. A sinalização via receptores que contêm gp130 indica que JAK 1 é uma quinase obrigatória e JAK 2 e TYK 2 servem de componentes adicionais.
Presume-se que a base precisa para a interdependência é a composição multicadeia e subunidades dos receptores. Esta teoria apoia-se no facto verificado de que os receptores do interferão α/β e interferão γ consistem de, pelo menos, 2 cadeias e foi demonstrado que a cadeia β do receptor do interferão α/β se liga a JAK 1, enquanto que TYK 2 se associa com a cadeia α. No caso do interferão γ foi mostrada a associação de JAK 1 com a cadeia α, enquanto que JAK 2 se associa com o complexo do ligando.


Tem-se tornado claro que as JAKs fosforilam múltiplos substractos, sendo os mais importantes destes substractos os próprios receptores.
Via de sinalização STAT
De entre os subtractos da JAK melhor caracterizados estão os STAT, e a activação de praticamente todos os receptores de citoquinas, interferãos e hormona de crescimento levam à activação de um ou mais STAT. Por exemplo, a activação de IL-3 das células hematopoiéticas leva à activação dos STAT-1, STAT-3, STAT-5 e STAT-6. A natureza dos STATs activados depende mais da linha celular em estudo do que da citoquina ou natureza do JAK activado pela interacção citoquina-receptor. Conclui-se daqui que a natureza do STAT activado por uma citoquina não depende nem do receptor da citoquina nem do JAK por si próprio.
STAT ( signal transducers and activators of transcription )
STAT é uma via de sinalização que se inicia no citoplasma mas rapidamente se faz a translocação para o núcleo, para activar a transcripção dos genes alvo. STAT joga um papel fundamental na via de transdução do sinal associada com várias citoquinas e neuroquinas, nas quais se incluem interleuquinas, interferãos, eritropoietina, prolactina, hormona de crescimento, oncostatina M ( OSM ) e factor neurotrófico ciliar ( CNTF ). Existem descritos na literatura um total de 8 STATs diferentes. O STAT 3 existe em 2 isoformas ( STAT 3A e STAT 3B ) e o mesmo sucede com o STAT 5 que apresenta também 2 isoformas ( STAT 5A e STAT 5B ) codificados por 2 genes diferentes, localizados no cromossoma 17, diferindo o STAT 5A do STAT 5B principalmente nos domínios terminais amino e carboxílico, com o domínio de transactivação apresentando as maiores divergências. Ambos os genes STAT 5 jogam um papel importante na proliferação das células hematopoiéticas induzida por IL-3, IL-5 e GM-CSF, bem como na proliferação do epitélio mamário induzida pela prolactina.

Estrutura de STAT
STAT apresenta uma estrutura com 5 domínios bem definidos, que incluem o domínio conservado N-terminal, domínio de ligação ao DNA, domínio putativo SH 3-like, um domínio SH 2 e um domínio de transactivação C-terminal. A região amino terminal parece ser crítica quanto à função do STAT, visto que pequenas delecções nesta região amino-terminal eliminam a capacidade do STAT ser fosforilado.
O domínio putativo SH 3-like tem ainda a sua função desconhecida mas o domínio SH 2 parece ser responsável importante na função sinalizadora do STAT. Este domínio SH 2 é crítico para o recrutamento dos STATs para os receptores activados, e é requerido para a interacção com JAK e Src quinases. O SH 2 é fundamental para a homodimerização e heterodimerização que, por sua vez, parece ser importante para a localização nuclear e ligação ao DNA.
Imediatamente a seguir ao domínio SH 2, por volta do resíduo 700, todos os STATs contêm um resíduo de tirosina, que joga um papel importante na activação STAT sendo que a fosforilação deste resíduo de tirosina é essencial para a activação e dimerização dos STATs.
O domínio C-terminal do STAT é requerido para transactivação transcripcional.
STATs que normalmente são de localização citoplasmática, são activados quando são fosforilados no resíduo tirosina que se situa cerca da posição 700, o que facilita a dimerização e translocação para o núcleo. A fosforilação das STATs ocorre imediatamente após a ligação dos factores de crescimento ou interferãos aos seus receptores.
As JAK quinases parecem não apresentar especificidade por uma STAT particular, uma vez que diferentes receptores activam uma STAT comum apesar de activarem distintivamente diferentes JAK quinases. A especificidade pela fosforilação dos STATs é determinada pelos locais de ancoragem para os STATs presentes no receptor e não nas JAK quinases
As 2 isoformas do STAT 5 ( STAT 5A e STAT 5B ) e o STAT 6 são activados pelos receptores IL-3, IL-5 e GM-CSF.
Duas vias independentes medeiam a fosforilação do JAK e STAT 3. Estes resultados sugerem que quinases da família Src medeiam a fosforilação do STAT 3 mediada pela interacção IL3-receptor e jogam um papel crítico na via da transdução do sinal associado com a proliferação das células mielóides. Isto sugere que a activação STAT 3, seguindo a estimulação das citoquinas de algumas células hematopoiéticas, pode seguir uma via diferente da que se verifica com STAT 5. Enquanto que uma ou ambas as isoformas STAT 5 actuam directamente com JAK 2, que por sua vez medeiam a fosforilação, a activação de STAT 3 pode requerer interacção com c-Src, a qual medeia a sua fosforilação.
Diferentes STATs podem ser fosforiladas por diferentes tirosina quinases sob condições diferentes. Tal como v-Src, v-Abl e BCR-ABL, foi demonstrado para STAT a transformação das células hematopoiéticas e torná-las citoquina-independentes para o crescimento.
BCR-ABL, constitutivamente, activa STAT 1 e STAT 5, com pouca ou nenhuma activação do JAK. Ao contrário do que sucede com as células B, transformadas pelo v-abl, hiperexpressão do domínio negativo dos mutantes JAK quinase nestas linhas celulares B não tem efeito na fosforilação STAT. Células Filadelfia positivas expressam STAT 5 activado constitutivamente e hiperexpressão do domínio negativo STAT 5 pode suprimir o estado transformado destas células. Activação de STAT pelo BCR-ABL é dependente da presença dos domínios SH2 e SH3, sugerindo interacção Src-like. Este facto dá uma base molecular para a incapacidade de v-Abl directamente activar STATs , uma vez que esta oncoproteina v-Abl não possui o domínio SH3, que está presente no oncogene BCR-ABL.
Há 2 vias independentes de mediação da activação do STAT, sendo uma dependente do JAK e uma independente das Src quinases.
Transdução do sinal, em resposta a diferentes ligandos, envolve a activação da via de sinalização Ras. Na via Ras, tipificada pela transdução do sinal em resposta a ligandos, como eritropoietina ou IL-3, a estimulação de citoquinas resulta no recrutamento e fosforilação da tirosina do Shc. Após a fosforilação, Grb2 associa-se com Shc e Sos, que leva a um aumento de GTP ligado ao Ras, activação do Raf-1, seguido pela activação de MAPK ( mitogen activated kinases ) e indução dos genes de resposta primária como c-myc, c-fos, etc.
Via de sinalização PI3K-akt
Angiogénese e proliferação celular tumoral: acção de VEGF e EGFR
Foi demonstrado que as células se fizeram equipar com “vias de segurança” para a eventualidade de mau funcionamento de uma via primária de sinalização. Esta eventualidade de vias de segurança permite as células sobreviverem à perda da via de sinalização primária. Assim, embora as vias Ras e JAK quinases medeiem sinais diferentes, está a evidenciar-se que pode haver interdependência entre as 2 vias. Fosforilação dos receptores JAK quinase cria potenciais locais de ancoragem para as moléculas adaptadoras como Src-quinase, subunidade p85 do PI3K, proteínas STAT e outras quinases como Src quinases que se podem associar a receptores fosforilados através dos domínios SH2. Uma vez recrutado ao complexo receptor, a JAK quinase pode fosforilar múltiplas cascatas de sinalização para proteger as vias ras e JAK quinase.
Truncagem no C-terminal da cadeia β do IL-3 resulta na perda da activação da via de sinalização ras, embora a capacidade para activar JAK 2 e induzir a mitogénese fosse mantida, dando evidência para a existência de uma via de sinalização alternativa da citoquina. JAK quinase alveja tanto uma via ras como uma via STAT para exercerem seus efeitos biológicos.
Activação da via ras, da via PI3K e das proteínas STAT, leva ao aumento da indução dos factores de transcripção, tais como c-jun.
Vários estudos reportaram o envolvimento da actividade JAK quinase na modulação da função PI3K.
A estimulação GM-CSF dos neutrófilos desencadeia a activação do JAK 2, STAT 3, STAT 5B e PI3K.
A maquinaria celular de transdução do sinal está programada para responder a alterações nos estímulos por integração de diversas vias de sinalização para gerarem uma resposta orquestrada.
Regulação negativa da actividade JAK
A participação das JAK quinases em múltiplas cascatas de transdução de sinal, em resposta a citoquinas e factores de crescimento, garante a necessidade de uma regulação negativa da actividade das JAK quinases. Este mecanismo de regulação negativo pode ser necessário para impedir a possibilidade da activação constitutiva da JAK quinase que pode levar a aberrante activação da cascata de sinalização e inapropriada expressão genética.
Tal cenário é validado pela observação de que a sinalização da JAK quinase foi implicada em situações patológicas, como leucemias e outras doenças malignas hematológicas.
Envolvimento da via de degradação mediada por proteossoma
Vias de degradação mediadas por proteossomas podem modular a actividade da JAK quinase, facto este corroborado pela observação de que a degradação de STAT e receptores das citoquinas e factores de crescimento também é vista na regulação da sinalização.
Inibidores dos proteossomas potenciam a activação, induzida por IL-2 e IL-3, da via de sinalização JAK quinase.
Regulação negativa pela PTP ( protein tyrosine phosphate )
PTP regulam a actividade quinase das tirosina quinases por desfosforilarem os resíduos tirosina envolvidos na função catalítica. Vários estudos demonstraram o papel das PTP na regulação da via de sinalização JAK quinase.
SHP-1, uma PTP, tem a capacidade de inibir a tirosina. Também foi comprovado um papel importante de SHP-2, outra PTP, na regulação negativa da JAK quinase nas células estimuladas pelo interferão.
Foi feita uma detalhada caracterização molecular de interacções específicas entre SHP-2 e JAK quinases, tendo sido demonstrado que SHP-2 é tirosina fosforilada pelo JAK 1 e JAK 2, mas não por JAK 3, através de associação directa. A associação SHP-2 e JAK quinase não requer o domínio SH2 do SHP-2 ou o domínio quinase-like nos JAKs.
Actividade fosfatase da SHP-2 parece não ser essencial para a interacção JAK-SHP-2 como se pode inferir pelo facto de tornar a fosfatase SHP-2 inactiva não ter feito abolir a interacção entre SHP-2 e JAK quinase.
A família das proteínas CIS/JAB/SOCS/SIS modula a actividade das vias de sinalização JAK.
.gif)
Estruturas de cada membro da família SOCS
Estas proteínas, induzidas por citoquinas, jogam um papel fundamental na regulação negativa dos sinais das citoquinas processadas pelas JAK quinases. Estas proteínas possuem domínios SH2 que permitem interacções proteína-proteína com membros da família dos receptores das citoquinas e outros componentes de sinalização. Estas proteínas são CIS ( proteínas contendo SH2 induzido por citoquinas ), SOCS ( supressores da sinalização das citoquinas ), SIS ( indutor STAT inibidor STAT ) e JAB ( proteína ligada a JAK ).
.jpg)
Proteínas SOCS/CIS e seus indutores
Proteínas SOCS são pequenas proteínas que possuem domínios SH2 e uma Box SOCS/CIS. A família SOCS, composta por 8 elementos, joga um papel crítico na modulação dos sinais propagados por diversos receptores de citoquinas.
CIS 1 foi o primeiro membro da família SOCS a ser identificado, e era uma resposta inicial para IL-2, IL-3 e eritropoietina, sendo associada com estes receptores.
SOCS 1 ( JAB ou SSI 1 ) foi identificado como potente inibidor das JAK quinases. A SOCS 1 interactua com o domínio quinase JAK 2 e suprime a via de transdução do sinal da IL-6.


Mecanismos moleculares de acção dos membros da família SOCS
SOCS 3 suprime a via de sinalização mediada pela eritropoietina através da ligação ao receptor da eritropoietina e JAK 2. Regiões N-terminal e C-terminal do domínio SH2 do SOCS 3 são necessárias para ligação de JAK 2 ao receptor da eritropoietina. Uma região citoplasmática do receptor da eritropoietina, que contem o Y401, é responsável pela ligação do SOCS 3 e é requerida para a actividade inibidora óptima do SOCS 3.
A actividade óptima das proteínas SOCS, e consequente inibição da actividade das citoquinas, requer as regiões SH2 e N-terminal das proteínas SOCS.
A ligação JAK/SOCS requer o domínio SH2 do SOCS, bem como os 12 amino-ácidos da região adjacente ao domínio SH2, que localiza 2 resíduos ( Ile 68 e Leu 75 ) que são conservados na proteína SOCS.
A família de proteínas CIS/JAB/SOCS/SIS regula a via de sinalização JAK por múltiplos mecanismos, sendo que algumas destas proteínas, como JAB e CIS 3, são capazes de ligar ao domínio quinase de JAK2 levando à inibição da actividade catalítica.

Via de transdução de sinal
JAK é também um mediador do sinal da apoptose. O JAK quinase medeia a resposta proliferativa de uma série de citoquinas e factores de crescimento, indicativa da indispensabilidade ao estímulo de crescimento total fornecido pela via da transdução do sinal das citoquinas e factores de crescimento. Isto sugere que a actividade da JAK quinase jogue um papel importante na prevenção da apoptose.



Sinalização fas

Sinalização TFN
A actividade da JAK quinase, conjuntamente com outras vias de sinalização, como ras, PI3K, MAPK, pode contribuir para o estímulo proliferativo total.
Proteínas da família Bcl-2 modulam os níveis de apoptose e sobrevivência celular, positiva ou negativamente, influenciando a morte celular e/ou sobrevivência celular. Assim, os membros da família Bcl-2, Bcl-XL e Mcl-1 inibem a apoptose, enquanto que Bax, Bad e Bak aceleram a apoptose. A via de sinalização JAK foi implicada na modulação da sobrevivência celular e apoptose por regular os membros da família Bcl-2. A via JAK quinase medeia o efector da regulação da morte celular Bcl-XL.
Implicações da actividade anormal da JAK em situações patológicas
Várias mutações dos membros da família JAK se associam a situações patológicas. Está neste caso, por exemplo, a associação da mutação da JAK 3 com o síndrome da imunodeficiência combinada grave. Activação aberrante da actividade da JAK quinase foi implicada em várias patologias malignas hematológicas, como LMC, LLA, etc e essas alterações da actividade ligam-se a translocações cromossómicas várias.
Uma mutação de JAK quinase pode causar anormalidades semelhantes clinicamente a leucemia.
A função das JAK quinases foi estudada no desenvolvimento das doenças. Verificou-se que células deficitárias na função JAK 1 apresentam sinalização deficitária através das vias de sinalização de múltiplos receptores de citoquinas, tais como a família gp130, família de receptores do interferão e receptores contendo cadeia γ.
Por outro lado, verifica-se que deficiência na função JAK 2 se associa a defeitos na eritropoiese, e nos casos de ausência de função JAK 2 há deficiente resposta de sinalização aos receptores das citoquinas IL-3, do interferão e receptor da cadeia única ( excepto receptor de G-CSF ).
Na presença de deficiência de JAK 3 verificam-se defeitos severos do desenvolvimento linfóide e aberrações da maturação das células B e activação dos linfócitos T, bem como no desenvolvimento dos linfócitos NK ( a deficiência de TYK 2 também leva a uma função defeituosa das células NK ). Mielopoiese apresenta-se desregulada quando se verifica ausência de JAK 3, com aumento de neutrófilos imaturos e monocitose de sangue periférico conjuntamente com esplenomegalia. Nestes casos de deficiência de JAK 3 verifica-se expansão das células da linhagem mielóide.
Em resumo, pode dizer-se que JAK 3 joga uma importante função no desenvolvimento das células das linhagens linfóide e mielóide.
Tal como interacções entre factor de crescimento e receptores, também interacções das citoquinas com os seus receptores levam à formação de múltiplos complexos proteicos como JAK, Src quinase, STAT, ras, MAPK, PI3K, PTP, CIS/JAB/SOCS.
Interessante verificar-se que receptores de citoquinas e interferãos diferentes interagem com várias moléculas de sinalização, tais como JAK, STAT, serina/treonina quinases, quinases duplas, fosfatases e seus reguladores negativos e ainda assim cada citoquina provoca respostas bioquímicas e biológicas distintas para cada célula.
É claro que a estoiquiometria dos complexos formados pela interacção citoquina-receptor joga um papel fundamental nas distintas respostas verificadas.
A via de sinalização JAK, e nomeadamente a mutação encontrada JAK2V617F constitui um grande avanço no conhecimento da patogenia das neoplasias mieloproliferativas, como sejam policitemia vera, trombocitemia essencial e mielofibrose primária. Esta descoberta de JAK2V617F traduziu-se numa revisão dos critérios de diagnóstico pela sua incorporação nesses critérios , assim como outras alterações como mutação no exon 12 de JAK ou do receptor da trombopoietina ( c-MPL ).
A presença ou não do JAK mutado nos doentes com neoplasias mieloproliferativas permitiu também estabelecer diferenças, sejam clínicas, evolutivas ou de prognóstico. Também o tratamento destas doenças sofreu grande evolução dado ter sido direccionado à molécula mutada JAK2V617F.
As proteinas quinase, JAK, são tirosina quinases que jogam um papel fundamental nas vias de transdução do sinal activadas por receptores celulares como resposta aos diferentes factores de crescimento ( eritropoietina, trombopoietina, GM-CSF ), interferãos e citoquinas. As proteínas da família JAK regulam, por meio dos receptores celulares específicos, a hematopoiese, a imunidade, a inflamação e o crescimento celular.
JAK 1 transmite sinais mediante diferentes receptores para interleucinas ( IL-6, IL-11, IL-2, IL-4, IL-7, IL-9, IL-15 e outras ), interferãos ( α, β e γ ) e factores estimulantes de colónias granulocíticas.
JAK 1 tem uma função principal na sinalização de diversas citoquinas pró-inflamatórias.
A activação de JAK 1 requer a presença de outros elementos da família, nomeadamente JAK 2, JAK 3 e TYK 2, para a activação dos receptores. JAK 2, ao contrário de JAK 1, pode fazer homodímeros, pelo que não precisa de outros JAKs para a transdução de sinal dos receptores activados de eritropoietina, trombopoietina e GM-CSF, mas por outro lado a JAK 2 precisa de TYK 2 para a activação dos receptores de interleucinas IL-12 e IL-23, e JAK 1 para os receptores de γ-interferão.
No que diz respeito às situações de neoplasias mieloproliferativas, é de salientar que os factores de crescimento eritropoietina, trombopoietina, GM-CSF, SCF ( stem cell factor ), IL-3 e IGF-1 ( insulin groth factor-1 ), a que os progenitores hematopoiéticos são hipersensíveis, utilizam JAK 2 como sinalizador celular. JAK 3 se expressa unicamente nos linfócitos. TIK 2, que heterodimeriza com JAK 1 e JAK 2, desempenha uma acção importante na resposta imune.
A mutação somática activante ( provocante de ganho de função ), JAK2V617F, produz-se no domínio autorregulador negativo, JH2, domínio este que na sua forma normal, inibe a função de JH1 mas que em mutação deixa de efectuar a sua função inibidora como estabilizador do domínio activo JH1 de JAK 2.
Em condições normais, os receptores dos factores de crescimento ou das citoquinas não apresentam actividade intrínseca, e esta actividade só se manifesta quando um ligando se une, ou o factor de crescimento ou uma citoquina, ao seu receptor específico. Na mutação de JAK2V617F, no domínio inibitório JH2, há activação da forma constitutiva da via de transdução do sinal. A activação constitutiva da via de sinalização, no caso da mutação JAK2V617F é continua, e não autorregulada, e como consequência desta actuação produz-se a fosforilação-activação da STAT que por sua vez induz a activação de múltiplas vias de sinalização como MAPK ou PI3K.
As STAT penetram no núcleo da célula e vão interferir nos processos de proliferação e diferenciação, regulação celular e apoptose, alterando o comportamento celular. A regulação negativa da via activada de sinalização JAK é mediada pelas proteínas SOCS ( supressor of cytokine signalling ) que se unem às JAKs e inibem a actividade catalítica. JAK2V617F pode fugir ao mecanismo de feedback do SOCS3.
O mecanismo sequencial normal fisiológico da sinalização é o seguinte:
factor de crescimento ou citoquina liga-se ao receptor, segue-se a activação
do receptor, fosforilação de JAK 2 e activação de STAT
do receptor, fosforilação de JAK 2 e activação de STAT
No caso da mutação JAK2V617F não se verifica a sequência fisiológica normal, já que independentemente do factor de crescimento ou citoquina, o estado de activação e transdução do sinal é permanente e escapa à normal regulação pela célula.
A mutação JAK2V617F não se encontra situada no local de união do ATP do JAK 2, o que explica que devido à falta de inibição selectiva de JAK 2 dos inibidores desta quinase ( JAK 2 ) , que são ATP-competitivos e se fixam aos locais de ligação do ATP, se produz inibição do JAK 2 também na variante não mutada, com a consequente repercursão da hemopoiese normal, originando uma potencial mielodepressão.
Os potenciais electrostáticos da superfície que rodeiam os locais de união ao ATP são diferentes nas diversas JAKs, o que pode constituir uma estratégia para maior selectividade.
A maioria dos inibidores ATP-competitivos de JAK 2 reduzem ou inibem a proliferação e sobrevivência celular de linhagens celulares e células de doentes com a mutação JAK2V617F. De referir que as células com a mutação são mais sensíveis aos inibidores JAK2 do que as células sem aquela mutação.
As citoquinas IL-6 e α-TNF estão relacionadas ao estado hipercatabólico e aos sintomas constitucionais, como fadiga e emagrecimento. A JAK 1 é a tirosina quinase envolvida na produção daquelas citocinas inflamatórias ( IL-6 e α-TNF ), citocinas que para além de se encontrarem em altas concentrações em estados inflamatórios, também se observam aumentadas na mielofibrose primária.
A inibição da JAK2V617F, STAT 5 e erk ½ resulta numa redução da proliferação celular e indução da apoptose.
JAK 2 tem como função normal provir instruções para a formação de uma proteína que promova o crescimento e divisão celular. Esta proteína é parte de uma via de sinalização e divisão celular, sendo a via de sinalização denominada de JAK/STAT, que tem por missão transmitir sinais químicos do exterior da célula para o núcleo celular. JAK 2 é crucial no controlo da produção das células sanguíneas a partir das stem cells hematopoiéticas que estão localizadas na medula óssea e têm potencial para se diferenciarem em eritrócitos, leucócitos ou plaquetas.
A mutação JAK2V617F é uma mutação somática, ou seja é adquirida durante a vida do doente e exprime-se apenas em algumas células e não noutras. A mutação JAK2V617F é a mais comum mutação somática e consiste da substituição do aminoácido Valina pelo amino-ácido Fenilalanina na posição 617 na proteína JAK2. Esta mutação JAK2V617F aparece em cerca de metade dos doentes com trombocitémia essencial, sendo que um pequeno número de doentes têm uma mutação somática noutra localização do gene JAK 2 conhecida por exon 12.
A mutação JAK2V617F encontra-se em cerca de 96% dos casos de policitémia vera a que se somam cerca de 3% de doentes com policitémia vera que apresentam mutação no exon 12 do gene JAK2.
A mutação JAK2V617F origina uma permanente activação da proteína JAK 2, resultando numa aumentada sobrevivência e produção dos eritrócitos.
No caso da mielofibrose primária a mutação JAK2V617F encontra-se em cerca de 50% dos doentes e uma pequena minoria destes doentes apresentam a mutação do exon 12. No caso da mielofibrose primária, a continua activação da proteína JAK 2 leva a uma hiperprodução de megacariócitos, que por sua vez estimulam outras células a libertarem colagéneo, o que vai originar a mielofibrose que posteriormente levará a uma medula óssea hipocelular.
O gene do JAK 2 está localizado no braço curto do cromossoma 9, na posição 24.
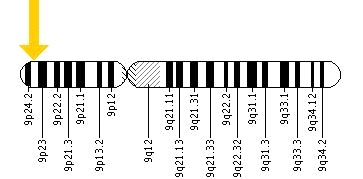
Cromossoma 9

Fusão BCR-ABL
Baseando-nos no conhecimento da mutação somática JAK2V617F, podemos considerar que as neoplasias mieloproliferativas, nomeadamente policitémia vera e mielofibrose primária, trombocitose essencial, representam, cada uma delas, 2 entidades patológicas diferentes, sendo uma positiva para a mutação e outra negativa desta mutação.
JAK 1 pode ser inibido por agentes patogénicos. É o caso de poliomavirus que ligando-se a JAK 1 inactiva o sinal através do receptor do interferão. Por outro lado, virus como o de Marburg, previne a fosforilação da tirosina de JAK 1 e subsequente fosforilação de STAT 1 e STAT 2.

Inactivação de JAK 1
JAK 1 está envolvido na via da transdução do interferão α/β e do interferão γ. A recíproca interdependência entre a actividade da JAK 1 e TYK 2 na via do interferão α, e entre JAK 1 e JAK 2 na via do interferão γ pode refletir uma necessidade destas quinases na correcta montagem dos complexos receptor-interferão.
JAK é uma família de proteínas tirosina quinase, não receptores, localizadas no citoplasma e não na superfície celular, que quando activadas estimulam uma cascata de reacções bioquímicas envolvidas na produção de citoquinas pró-inflamatórias. Hiperactivação do JAK pode levar a inflamação e destruição tecidular. Cada JAK tem especificidade por um diferente grupo de receptores das citoquinas. A função das proteínas JAK está ligada à função das citoquinas que se ligam aos receptores envolvidos.


Dimerização
JAK não funciona sózinho, sendo que cada receptor de citoquinas requer, pelo menos, 2 JAKs associados. Os pares de JAKs podem ser homodímeros ou heterodímeros.
As citoquinas que utilizam a via de sinalização JAK têm sido divididas em 2 categorias:
- Tipo I : incluem interleuquinas, receptores de citoquinas tanto cadeia γ como não cadeia γ, e alguns factores hematopoiéticos
- Tipo II: incluem interferãos ( α, β ou γ ) e IL-10
Janus quinase é uma, de entre 10 famílias conhecidas, de tirosina quinases não receptores.
Cada um dos membros da família JAK tem um domínio quinase e um domínio pseudo-catalítico, e ambos estes domínios ligam receptores de citoquinas através do domínio N-terminal FERM ( Band 4.1, Ezrin, Radoxin, Moesin ).
Depois de se ligarem as citoquinas aos seus receptores, JAKs são activados e fosforilados os receptores, criando locais de ancoragem para moléculas de sinalização, especialmente STAT.
Mecanismo de actuação do sistema JAK/STAT
.jpg)
.jpg)
Figura 1
.jpg)
.jpg)
Figura 2
.jpg)
Figura 3
.jpg)
Figura 4
.jpg)
Figura 5
.jpg)
Figura 6
.jpg)
Figura 7
JAK é formado por um JH1, JH2, um domínio SH2-like composto por JH3, JH4 e JH5, e um domínio FERM no N-terminal composto por JH6 e JH7. O FERM está implicado na mediação das interacções com proteínas transmembrana, tais como os receptores das citoquinas. JAK é, para algumas citoquinas mas não para todas, importante na regulação da expressão da superfície celular dos receptores cognate ( cognate são 2 biomoléculas que tipicamente interagem, como por exemplo um receptor e seu ligando ). O FERM liga o domínio quinase e regula a actividade catalítica positivamente.

gp130
A subunidade gp130 do receptor tem o JAK 1 como essencial, e relaciona-se com diversas citoquinas, como IL-6, IL-11, oncostatina M, factor inibitório de leucemia ( LIF ), factor neurotrópico ciliar ( CNF ), bem como G-CSF e interferãos.
O JAK 2 é essencial para transdução do sinal via receptores tipo I, como citoquinas hormona-like como a leptina, hormona de crescimento, prolactina, eritropoietina, trombopoietina e, como transdutores do sinal dos receptores do tipo II, uma família de citoquinas que sinalizam através do receptor IL-3 ( IL-3, IL-5 e GM-CSF ). Também o JAK 2 é importante para as citoquinas que usam receptor gp130 e para alguns interferãos.
TYK 2 é essencial para a sinalização de interferãos, mas também para sinalização do IL-12, mas não para a sinalização por interferão α/β ou citoquinas que usem gp130.
Deficiência em JAK 2 leva a morte embrionária precoce por falência da eritropoiese.
A família JAK, com cada constituinte envolvido em graus diferentes que podem ir de total comprometimento a ausência total desse comprometimento, está envolvido no crescimento celular, sobrevivência, desenvolvimento e diferenciação das células, mas é criticamente importante, esse envolvimento, nas células hemopoiéticas e imunológicas. Completa ausência de função de JAK 1 ou JAK 2 é incompatível com a vida. Pelo contrário, ganho de função de cada uma das JAKs se associa com transformação maligna, sendo a mais comum a associação com doenças mieloproliferativas.
O genoma humano documenta a existência de 518 proteínas quinases e 90 membros da família quinase formam o grupo das proteínas tirosina quinase ( PTK ) que são as enzimas responsáveis pela transferência de γ fosfato dum nucleotídeo purina trifosfato ( ATP, GTP ) para grupos hidroxílicos de resíduos tirosina específicos de outros substractos proteicos.
A fosforilação da tirosina estimula a actividade enzimática, mas também permite o recrutamento das proteínas de sinalização da cascata pela alteração da conformação e criando locais de ligação a proteínas com domínios Src homologia 2 ( SH 2 ).
As PTK dividem-se em 2 classes, dependendo da sua localização celular.
Depois de se ligarem ao seu ligando específico, o receptor estimulado activa o PTK citoplasmático associado e a fosforilação da tirosina subsequentemente recruta adicionais proteínas de sinalização por processos locais de ligação.
Eficaz resposta, inata e adaptativa, requer sinalização JAK funcional para a protecção do organismo de infecções ou tumores e mutações, que levem à perda de função, origina algumas das mais comuns imunodeficiências graves. Pelo contrário, mutações activantes ou que levem a perda funcional de membros da família JAK, originam transformações malignas das células linfocíticas ou mielóides.
Mutações do domínio FERM foram associadas a alterações de actividade quinase. Em contraste com o seu papel na actividade quinase, o domínio FERM parece ser essencial para a ligação do JAK ao receptor.
O domínio FERM medeia a interacção do JAK com o seu receptor de citoquinas transmembrana cognate, e regula a actividade quinase.
Os receptores de citoquinas tipo I ou tipo II dependem da transmissão de sinal feita pelas PTKs, como JAKs, visto não terem actividade quinase ligada aos receptores. Um JAK, homodimérico ou heterodimérico, selectivamente se associa com diferentes receptores de citoquinas para transmitirem o sinal após activação de receptor específico.
A heterodimerização TYK 2 com JAK 2, não só é necessária para a sinalização do γ-interferão, como é precisa para a diferenciação das células Th1 produtoras de γ-interferão a partir de células Th naif. IL-12 é a citoquina responsável pela geração de Th1. O receptor de IL-12 está associado a TYK 2 e JAK 2, e activa principalmente o factor transcripcional STAT 4.
IL-12 e IL-23 sinalizam através de TYK 2 e JAK 2, mas enquanto a activação de TYK 2 dependente de STAT 4 é o principal evento sinalizador após estimulação por IL-12, a fosforilação do STAT 3 mediada por JAK 2 é a predominante quando a IL-23 é a envolvida.
IL-12 + receptor -----------> TYK 2 ( STAT 4 )
IL-23 + receptor -----------> JAK 2 ( STAT 3 )
Desta forma parece TIK 2 ser indispensável para o eixo interferão/Th1 e para a resposta imune mediada por IL-17/Th17 que é promovida pela IL-23.
Consequências de deficiência de TYK 2
A sinalização IL-12 é fortemente alterada na ausência de TYK 2, a cascata de activação STAT 3 e STAT 4 claramente reduzida, resultando na incapacidade de produção pelas células T de γ-interferão. A sinalização de interferão também depende da expressão funcional de TYK 2, e verifica-se uma maior susceptibilidade a infecções víricas e bacterianas nos indivíduos deficientes em TYK 2, dado que os eventos usando α-interferão, na sua sinalização, estão afectados. Esta alteração de TYK 2 leva a uma alteração de desenvolvimento da linhagem Th1, e a linhagem Th17 também parece ser afectada na ausência de TYK 2.

Vias de sinalização activadas por interferão α/β, interferão γ e interferão λ
TYK 2 também é importante, para além da sinalização das células Th, na sinalização das células dendríticas.
Resposta imune inata executada pelas células NK ( natural killer ) é afectada nas deficiências de TYK 2. IL-18 actua sinergisticamente com IL-12 para activar as células NK.
A ausência de TYK 2 foi identificada até hoje em apenas 1 doente, e a sintomatologia apresentada era de múltiplas infecções oportunísticas de vários órgãos por virus, bactérias e fungos. Ausência de TYK 2 nas células T, interfere com múltiplas vias de sinalização de citoquinas. Activação de STAT 1, STAT 2, STAT 3 e STAT 4, após estimulação com interferão tipo I, estava ausente. Quando a estimulação celular era feita por IL-12 ou IL-23, a activação de STAT 4 não se verificava.
A ausência de TYK 2 associa-se a uma deficitária resposta Th1 e aumentada resposta Th2.
Consequências da deficiência de JAK 3
Os 4 membros da família JAK estão envolvidos, de forma crucial, na transmissão do sinal das citoquinas, que é responsável pelo desenvolvimento, diferenciação e proliferação das células T, mas, no entanto, apenas JAK 3 está envolvido em todas estas funções. A deficiência de JAK 3 leva a deficiente desenvolvimento e proliferação linfocitária. O desenvolvimento dos linfócitos T e B, a partir de um progenitor hemopoiético, é estritamente dependente de IL-7 ( outras citoquinas importantes na proliferação e sobrevivência dos linfócitos são IL-2, IL-9 e IL-15; IL-4 e IL-21 são importantes na regulação da função B, incluindo a produção de imunoglobulinas ).
Na ausência de JAK 3, não pode haver sinalização IL-2 nem activação STAT 5.
Sinalização JAK 3 influencia, não só o desenvolvimento e sobrevivência das células T, como também a diferenciação celular Th, sendo crucial na diferenciação Th2, sendo IL-4 a mais crítica citoquina interveniente nesta diferenciação e IL-4 depende de JAK 3 para transmitir o sinal. Assim, na ausência de JAK 3, IL-4 origina a activação de STAT 6, com consequente bloqueio da diferenciação celular Th2. Parece que a ausência de JAK 3 também influencia negativamente a diferenciação Th1 por modificações epigenéticas.
Deficiência em JAK 1
Esta ausência ainda não foi descrita, provavelmente porque é incompatível com a vida devido a déficits neurológicos graves. Na deficiência de JAK 1 pode haver importante alteração da linfopoiese e falência a responder a sinais dos receptores classe II, receptores γc citoquinas e receptores de citoquinas que contenham subunidade gp130.
Deficiência de JAK 2
Tal como a ausência de JAK 1, também a ausência de JAK 2 é letal, sendo esta devido a eritropoiese deficitária. Estimulação do receptor da eritropoietina induz a fosforilação tirosina de JAK 2, requerida para a actividade biológica da eritropoietina. Perturbação da sinalização da eritropoietina e sua transdução, bem como de outras hormonas necessitantes de JAK 2 funcional, parece ser responsável pela eritropoiese que não se processa, e consequente morte fetal.
Mutação JAK 2 em doenças malignas

A família JAK é essencial para a sinalização dos receptores das citoquinas durante a diferenciação das respostas imunológicas, mas também tem acção fundamental na mediação dos sinais no crescimento, proliferação e formação das células hematopoiéticas e de outros tecidos.
Uma importante associação da mutação JAK 2 é com neoplasias mieloproliferativas, grupo de doenças hematológicas onde se verifica um excesso de proliferação de uma ou mais linhagens celulares mielóides.
Leucemia mielóide crónica foi associada a um oncogene específico, o BCR-ABL 1, causado pela translocação do cromossoma 9 e 22 ( cromossoma Filadélfia ). Pelo contrário, as outras neoplasias mieloproliferativas não se associam à ABL quinase, mas ligam-se a uma mutação, V617F, que resulta numa quinase constitutiva activa, que se autofosforila, e se torna espontaneamente activa, tornando as células hematopoiéticas independentes das citoquinas para o crescimento e sobrevivência.

Mutações com ganho de actividade de JAK 3
Mutação JAK 2 é frequentemente encontrada nas neoplasias mieloproliferativas, mas também já foram descritas mutações JAK 2 em doentes com LLA, LMA e leucemia megacarioblástica aguda. Os pontos de mutação, como o V617F no JAK 2, resultam em fosforilação tirosina constitutiva de JAK 2 e promovem a sobrevivência e proliferação celulares, independentes de sinais recebidos por citoquinas, que se ligam ou não aos seus receptores. Mutações JAK 3, activação constitutiva JAK 3 e fosforilação da cascata STAT 5, relacionam-se com a leucemia megacarioblástica aguda. Também LMA já foi relacionada com mutação JAK 1, enquanto que mutação TYK 2 ainda não foi referida como associada a doenças malignas hematológicas. Mutações do JAK 3, para além de doenças malignas hematológicas, já foram reportadas a tumores sólidos, nomeadamente da mama e do estômago.
Após a ligação das citoquinas aos seus receptores, JAKs são activados e fosforilam os receptores, criando locais de ligação para as moléculas sinalizadoras, especialmente os STAT.

O sistema JAK/STAT é composto por 3 elementos principais: receptor, JAK e STAT, sendo este último o que leva o sinal ao núcleo e DNA.
Depois da citoquina se ligar ao receptor, o JAK adiciona um fosfato ao receptor. Isto atrai o STAT, que também é fosforilado, e liga-se a outro STAT, formando um dímero. Este dímero move-se para o núcleo, onde se liga ao DNA, e origina a transcripção dos genes. As enzimas que adicionam os grupos fosfato são chamadas de proteínas quinases ( PTK ).
Cascata MAPK
Dado os membros das famílias de receptores de citoquinas tipo I ou tipo II não possuirem actividade catalítica quinase, eles transferem essa acção para a família JAK, para fosforilarem e activarem as proteínas da cascata envolvidas na sua via de transdução de sinal. Os receptores existem como polipeptídeos emparelhados, que exibem 2 domínios intracelulares de transdução de sinal. JAK associa-se a uma região rica em resíduos de prolina, no domínio intracelular, adjacente à membrana celular, e chamada de Box1/Box2. Após o receptor se associar ao seu respectivo ligando ou citoquina, sofre uma alteração conformacional, levando os 2 JAKs próximos, o suficiente, para se fosforilarem um ao outro. A autofosforilação dos JAKs induz uma alteração conformacional nele próprio, permitindo fazer a transdução do sinal intracelular através de outras fosforilações e activando factores de transdução chamados STAT. O STAT activado, dissocia-se do receptor e forma dímeros antes de os translocar para o núcleo onde regula a transcripção dos genes seleccionados
Sinalização cAMP
Mutação JAK2V617F e as neoplasias mieloproliferativas
As neoplasias mieloproliferativas se originam por proliferação clonal de um progenitor hematológico pluripotencial, levando à hemopoiese exacerbada com expansão de uma ou mais linhagens.
Os progenitores hematopoiéticos dos doentes com neoplasia mielopropliferativa apresentam hipersensibilidade a diversas citoquinas como eritropoietina, IL-3, stem cell factor, IGF-1, GM-CSF e trombopoietina.
Foram associadas várias mutações a diversas neoplasias mieloproliferativas, como BCR-ABL 1 com LMC, BCR-JAK 2 com LMC atípica ou LMC BCR-ABL negativa, ou LMMC com Tel-PDGFRβ.

Mutação JAK2V617F: mecanismo de acção e inibidores de JAK2
A mutação V617F, trata-se de uma mutação pontual, consistindo de uma substituição de uma guanina por uma tiamina ( G → T ) no exon 14 do gene JAK 2, levando à substituição de uma valina por uma fenilalanina, na posição 617 da proteína codificada JAK2V617F.
No gene JAK 2 guanina substituída por tiamina
Na proteína JAK2V617F valina substituída por fenilalanina
.jpg)
Mutação JAK2V617F
Esta alteração é uma mutação somática detectada nas células eritróides e mielóides, mas não em células T ou da mucosa oral.

Mutação V617F e em exon 12
Relativamente à actividade tirosina quinase e sinalização intracelular induzidas pela mutação JAK2V617F, foi demonstrada autofosforilação constitutiva de JAK 2, fosforilação constitutiva do factor de transcripção STAT 5 e activação das vias ERK, PI3K, Akt, além da indução de transcripção dependente de STAT 5 na ausência de eritropoietina. Esta sinalização, exacerbada, leva à hipersensibilidade e à independência de factores de crescimento.
Citoquinas regulam muitos processos biológicos, incluindo respostas inflamatórias e imunológicas. O complexo equilíbrio das citoquinas, balanceia os efeitos pró-inflamatórios e anti-inflamatórios, tentando igualizá-los. Um desequilíbrio entre as citoquinas pró-inflamatórias e anti-inflamatórias pode resultar numa doença inflamatória.


Citoquinas pró-inflamatórias e anti-inflamatórias
A maioria dos receptores de citoquinas, que modulam a actividade hematopoiética, pertencem ao tipo I. Os receptores do tipo II são principalmente ligados a interferãos.
Interessante o facto de que a mutação JAK2V617F aparece em todas as neoplasias mieloproliferativas ( policitémia vera, trombocitose essencial e mielofibrose primária mas não na LMC ) mas a mutação no exon 12 do gene JAK 2 aparece apenas na policitémia vera.
Quando a mutação JAK2V617F é expressada nas células hematopoiéticas, várias vias de sinalização importantes para a proliferação e sobrevivência celulares são activadas, incluindo STAT 3, STAT 5, MAPK, erk ( extracelular signal-regulated kinase ) e PI3K-akt ( fosfoinositide 3 quinase ).
As mutações no gene JAK 2, exon 12 e JAK2V617F, mutuamente se excluem.
Os resíduos afectados pela mutação no exon 12 não estão estritamente localizados no domínio JH2, mas podem, ainda assim, contribuir para as interacções auto-inibidoras intradomínio.

Mutação exon 12
Expressão de JAK 1 nas células cancerosas, permite as células, individualmente, contrairem-se, potencialmente permitindo-lhes escapar ao tumor de onde pertencem e, assim, metastizarem para outros locais do organismo.
Sem comentários:
Enviar um comentário