Sinalizador
celular – Ubiquitina

Ubiquitina
A ubiquitina é uma proteína fundamental na regulação de processos biológicos, assinalando algum processo que deve acontecer ao substracto com o qual está ligado, como por exemplo as proteínas alvo serem degradadas pelo proteossoma.
O
proteossoma é uma proteína com função de degradação de outros
substractos, como um triturador que as células possuem.
http://youtu.be/93in7Bxo5W0
Cerca
de 600 genes codificam ubiquitina-ligases no genoma humano. Foi
descrita uma ubiquitina-ligase que controla a dinâmica mitocondrial,
denominada MULAN ( Mitochondrial Ubiquitin Ligase Activation of NF-kB
).
O
gene BRAC1, ligado ao cancro da mama e outros cancros, como pâncreas
e próstata, quando mutado pode eliminar a função
ubiquitina-ligase.
cDNA
ou DNA complementar é o DNA sintetizado por uma molécula de RNA
mensageiro numa reacção catalizada pela transcriptase reversa que
quando inserido numa célula, este cDNA pode levar ao aumento dos
níveis da proteína codificada.
RNAi
ou RNA de interferência é uma técnica que permite inibir a acção
dum gene escolhido, por exemplo o RNAi pode suprimir genes que causam
tumores diminuido o desenvolvimento do cancro.
A via da ubiquitinação tem 3 etapas, sendo que cada uma envolve uma enzima: E1 ( enzima activadora da ubiquitina ), E2 ( enzima conjugadora ) e E3 ( ubiquitina-ligase ).
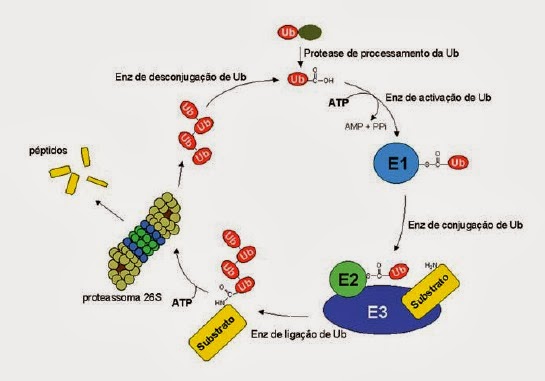
Via
ubiquitina-proteossoma
A via da ubiquitina-proteossoma é uma via proteolítica que, ao contrário das demais, consome ATP e inicia-se com a ubiquitinação da proteína alvo.
Os
alvos da ubiquitina são:
- receptores celulares da membrana celular
- moduladores de crescimento e supressores de tumor
- activadores e inibidores de transcripção
- reguladores de ciclo celular
- proteínas mutantes ou danificadas, sendo estas especialmente frequentes chegando a 30% das proteínas recém-sintetizadas
http://youtu.be/4DMqnfrzpKg
As chaperonas desempenham um papel activo no reconhecimento das proteínas com síntese defeituosa. As chaperonas são proteínas que auxiliam outras proteínas a se dobrarem, necessitando várias chaperonas de ATP, e algumas proteínas para atingirem a estrutura quaternária são dependentes das chaperonas. O reconhecimento da má dobragem das proteínas, ainda que não totalmente ilucidado, sabe-se que utiliza os grupos hidrofóbicos para esse reconhecimento.
As
chaperonas interferem activamente na ubiquitinação, interagindo com
as enzimas E2, pensando-se que as chaperonas são responsáveis pelo
reconhecimento e marcação da proteína pela ubiquitina.
Outro sistema importante para o reconhecimento de proteínas mal formadas é o ERAD ( Endoplasmic Reticulum Associated protein Degradation ) que é o responsável pela degradação das proteínas sintetizadas no retículo endoplasmático e que também utiliza as chaperonas como auxiliares. Estas proteínas após a ubiquitinação vão para o citoplasma e são degradadas pelos proteossomas.
A
via ubiquitina-proteossoma envolve várias etapas, nomeadamente:
- Activação: a enzima E1 activa a ubiquitina cujo resíduo C-terminal se liga covalentemente a um resíduo sulfidril-cisteína da E1 na presença de Mg²+ e com consumo de uma molécula de ATP que se degrada em AMP e Pi ( pirofosfato )
- Transferência: a ubiquitina é transferida para a enzima E2 e liberta-se a enzima E1
- Reconhecimento: a enzima E3 reconhece e liga-se à proteína alvo formando um complexo não covalente
- Ubiquitinação: o complexo E2-ubiquitina liga-se a E3 por forma à ubiquitina ser transferida de E2 para o grupo amina dum resíduo lisina da proteína alvo
- Libertação: a enzima E3 solta-se, libertando E2 e a proteína ubiquitinada
- Poli-ubiquitinação: as etapas 3, 4 e 5 repetem-se formando uma ou mais cadeias de ubiquitina, cadeias estas que se ligam aos resíduos de lisina da última cadeia
- Desubiquitinação: a cadeia de ubiquitina é reconhecida pelo proteossoma, sendo a proteína desubiquitinizada por enzimas próprias e a proteína desdobrada com consumo de ATP e hidrolisada, dando origem a pequenos peptídeos de 7-9 aminoácidos.
Apenas
proteínas que sofreram poli-ubiquitinação são capazes de acessar
ao núcleo proteolítico do proteossoma.
A
enzima E3 ( ligase ) é a responsável pela especificidade da
ubiquitinação capaz de fazer o reconhecimento e etiquetagem de
vários substractos.
A
ubiquitinação é um processo reversível em que as desubiquitinases
( enzimas hidrolíticas ) competem com o proteossoma, removendo as
ubiquitinas terminais da cadeia ou mesmo a cadeia inteira.
Ao
contrário da ubiquitinação, que é um processo reversível, a
proteólise não o é, uma vez que as proteínas são degradadas a
oligopeptídeos. Esta assimetria reaccional permite impôr uma
direcção ao sistema, permitindo por exemplo o ciclo celular.

Via
da ubiquitina-proteossoma ( UPP )
A ubiquitina é uma proteína das células eucarióticas, formada por 76 aminoácidos, e que tem uma função importante na regulação da síntese proteica, marcando as proteínas mal dobradas para estas serem degradadas pelos proteossomas.
A
ubiquitina, para além do processo proteolítico não lisossomal, das
proteínas mal dobradas, também tem acção não proteolítica no
transporte pela membrana, na estrutura e transcripção da cromatina,
na reparação do DNA e noutras vias sinalizadoras.
As proteínas encontram-se num estado dinâmico, sendo continuamente sintetizadas e degradadas, sendo o controlo dessa actividade e degradação vital para a célula. As vias proteolíticas são os meios pelos quais as células se libertam das proteínas indesejadas.
As
principais vias proteolíticas são a via lisossomal, via dependente
do cálcio e via da ubiquitina-proteossoma, sendo que esta última
difere das anteriores por ser selectiva e consumir ATP.
A
ubiquitina é uma molécula que tanto se encontra no núcleo como no
citoplasma; apresenta grande tolerância ao pH e à temperatura,
apenas variando entre a conformação dobrada e não dobrada,
justificada esta conformação por ser determinada por ligações de
hidrogéneo.
A
estrutura secundária da ubiquitina é formada por 3 segmentos em
α-hélice e 5 em folhas
β, tendo um peso
molecular de 8564 Daltons

Estrutura
tridimensional da ubiquitina
Ubiquitinas podem formar cadeias através da ligação de várias ubiquitinas, ocorrendo a ligação entre a glicina terminal e a lisina, podendo esta ligação ser feita várias vezes, originando di-, tri- ou tetra-ubiquitinas. A síntese da ubiquitina realiza-se nos ribossomas.
Tanto
a ubiquitina como sua via proteolítica participam de diversos
processos celulares. A ubiquitina liga-se à ciclina, na fase G1 da
mitose, ajudando a regular o ciclo celular; a ubiquitina é
responsável por estimular a diferenciação entre os tipos de
linfócitos B e T, e seus fragmentos funcionam extracelularmente como
imunossupressores. A ubiquitina tem acção na biogénese dos
ribossomas, modulação de receptores celulares, expressão génica,
reparação de DNA, resposta ao stress, morfogénese neuronal,
memória de longo prazo, ritmos circadianos e em diversas doenças
como Alzheimer, Down, cancro, HIV e Ébola.
A
ubiquitinação realiza-se com a ajuda de 3 tipos de enzimas: E1, E2
e E3. Só é conhecida uma E1 ( enzima activadora da ubiquitina ) que
transfere ubiquitina a todas as E2, sendo que a E2 já tem alguma
especificidade ao substracto, embora sejam as E3 as que apresentam
grande especificidade. Enquanto há apenas uma E1, existem 5-12
proteínas E2. As E3, de 250 kDa, são as responsáveis pelo
reconhecimento da proteína e transferência da ubiquitina de E2 para
E3. Actualmente aceita-se que a poli-ubiquitinação é mediada pela
enzima E4, que coopera com as E3 para estabelecer a cadeia de
poli-ubiquitina.
Todas
as proteinas intracelulares e muitas extracelulares estão
constantemente a ser reconstituídas, isto é são hidrolizadas nos
seus aminoácidos constituintes e novamente sintetizadas. Esta
situação de construção-destruição das proteínas é importante
para funções homeostáticas. A degradação das proteínas não é
feita para todas à mesma velocidade, indo o tempo de degradação
proteica de fracções de segundo até semanas ( como a actina e
miosina ) ou meses ( hemoglobina ). As células possuem múltiplos
sistemas proteolíticos usados na degradação das proteínas e
complexos mecanismos reguladores que assegurem os processos
proteolíticos contínuos são altamente selectivos.
A
maioria das proteínas intracelulares são degradadas pela via
ubiquitina-proteossoma. Algumas proteínas de superfície e as
proteínas extracelulares são tomadas por endocitose e degradadas
nos lisossomas que contêm várias proteases e hidrolases. Algumas
proteínas citosólicas são degradadas nos lisossomas após sofrerem
autofagia e sendo assim englobadas em vacúolos que se fundem com
lisossomas. Este processo é acelerado, nas células, pela falta de
insulina ou aminoácidos essenciais e, no fígado, pelo glucagon.
Outra
família importante de proteases citosólicas são as caspases que
fazem a clivagem nos resíduos do ácido aspártico. As caspases,
proteases de cisteina, têm uma função crítica na destruição dos
componentes celulares no processo de apoptose.
A
via ubiquitina-proteossoma liga a ubiquitina às proteínas marcadas
para degradação. Este processo de marcação leva as proteínas a
serem reconhecidas pela proteossoma que degrada as proteínas
ubiquitinadas a pequenos polipeptídeos.
A
via ubiquitina-proteossoma utiliza 3 enzimas denominadas E1, E2 e E3.
Enquanto E1 e E2 preparam a ubiquitina para a conjugação, a E3 (
ligase ) reconhece o substracto proteico específico e cataliza a
transferência da ubiquitina activada para esse substracto proteico.
Factores
de transcripção podem ser regulados por alterações na sua
localização intracelular. Um exemplo é o que se passa com NF-kB
activador transcripcional pro-inflamatório mantido fora do núcleo
pela sua interacção com IkB. A fosforilação desta molécula IkB
desencadeia a sua degradação, o que leva a que seja reconhecido
pela E3 β-transducina repeat
containing protein ( β-TRCP ). A IkB é ubiquitinada e degradada
rapidamente, permitindo que NF-kB fique livre e se dirija para o
núcleo. Este passo é de fundamental importância na aceleração do
processo inflamatório.
A
via da ubiquitina-proteossoma elimina selectivamente as proteínas
defeituosas, seja esse defeito por mutação, erro de síntese ou
lesão por radicais de oxigéneo ou desnaturação pelas altas
temperaturas.
A
via proteolítica extralisossomal dependente de ATP,
ubiquitina-proteossoma, é extremamente importante em diversos
mecanismos reguladores celulares como transdução de sinal, controlo
de ciclo celular, endocitose mediada por receptores, transcripção
genética, controlo de qualidade de síntese proteica, fornecimento
de aminoácidos em situação de carência nutricional ou condições
patológicas, apresentação de antigenes e apoptose.
O
sistema proteolítico lisossomal foi o primeiro sistema proteolítico
intracelular identificado, mas foram descobertos outros sistemas
intracelulares extralisossomais proteolíticos

Proteólise
lisossomal
A via da ubiquitina-proteossoma é uma via proteolítica, de fundamental importância para a viabilidade celular, dependente do ATP. A ubiquitina é formada por uma protease de 1000-1500 kDa, formada por subunidades de 34-110 kDa. É formada por um núcleo catalítico cilindríco, denominado proteosoma 20 S e uma porção 19 S onde se localizam as ATPases responsáveis pelo desenovelamento das proteínas a serem degradadas pelas actividades proteolíticas da subunidade 20 S.
A
ubiquitinação proteica é uma modificação pós-transducional
consistindo esta da adição de uma ou mais moléculas de ubiquitina
a proteínas específicas que alteram a função, localização
celular ou a direccionam para a degradação no proteosoma.

Proteosoma
Nas células eucarióticas, a principal via de degradação proteica de semi-vida curta é dependente do ATP e da ubiquitina e não uma via lisossómica. Na via ubiquitina-proteossoma são degradadas proteínas reguladoras de certas funções que por isso existem em curtos períodos de tempo, bem como proteínas danificadas ou com erros na sua síntese, que são marcadas para degradação por meio da ligase de cadeia poli-ubiquitina. Estas proteínas assim marcadas são reconhecidas e degradadas pelo proteosoma 20 S após terem sido desenoveladas no proteossoma 19 S. Esta via ubiquitina-proteossoma funciona no citoplasma e núcleo das células. Proteínas associadas ao retículo endoplasmático também são degradadas pela via ubiquitina-proteossoma após o seu retrotransporte para o citoplasma.
A
falta de apertado controlo do ciclo de divisão celular está
implicado no desenvolvimento tumoral. A via ubiquitina-proteossoma
joga uma função fundamental pelo menos em 3 períodos do ciclo de
divisão celular. Na transição G1/S, a iniciação da replicação
do DNA é desencadeada pela degradação dum inibidor duma cinase
dependente da ciclina. Quando este inibidor é fosforilado, uma E3 o
reconhece como substracto e dá-se a ubiquitinação e subsequente
degradação. Na mitose, a securina ( endoprotease envolvida na
separação dos cromossomas ) é ubiquitinada e degradada pela
proteossoma. Na fase final da mitose, as ciclinas e factores que
controlam a desmontagem do fuso mitótico sofrem degradação pela
via ubiquitina-proteossoma.
Esta
via ubiquitina-proteossoma também produz peptídeos antigénicos a
partir de proteínas de bactérias ou virus. Os peptídeos
apresentados às células citotóxicas T nas moléculas do complexo
major de histocompatibilidade da classe I resultam de proteólise,
limitada dos antigéneos, mediada pelos proteossomas. O γ-interferão
estimula este processo induzindo a troca de subunidades catalíticas
do proteosoma 20 S. O sistema dependente da via
ubiquitina-proteossoma degrada tanto as proteínas celulares ( self )
como as estranhas ( non-self ).
Os
peptídeos são apresentados às células citotóxicas T mas apenas
os derivados das proteínas estranhas desencadeiam resposta das
células T.
Sem comentários:
Enviar um comentário